Corrales-Guerrero L, Mariscal V, Nürnberg DJ, Elhai J, Mullineaux CW, Flores E, Herreroa A (2014). Subcellular Localization and Clues for the Function of the HetN Factor Influencing Heterocyst Distribution in Anabaena sp. Strain PCC 7120. J Bacteriol 196:3452-3460.
PatS and HetN both suppress heterocyst differentiation and seem to fulfill the specifications of the S component of a Turing-like model, but superficially, the two proteins seem very more different. PatS protein is 11 to 17 amino acids long while HetN is much larger, 287 amino acids long. HetN is part a large gene family of short chain dehydrogenases, while PatS is found only in a subset of cyanobacteria (Elhai & Khudyakov, 2018). What they have in common is a conserved, internal RGSGR amino acid motif, which in HetN is confined to a small cluster of related cyanobacteria (Figure 1).
All of the HetN sequence is dispensible for its function to regulate heterocyst differentiation, except for two regions: (1) the RGSGR motif, and (2) the N-terminal 27 amino acids, which may function as either a signal sequence (for export out of the cytoplasm) or a membrane anchor. The protein is expressed at the cell poles of heterocysts, near contacts with adjacent cells.
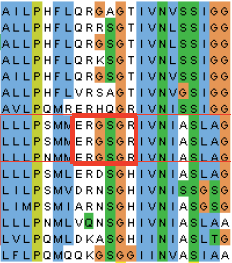
Figure 1. See text for explanation. Taken from Corrales-Guerrero (2014).
Elhai J, Khudyakov I (2018). Ancient association of cyanobacterial multicellularity with the regulator HetR and an RGSGR pentapeptide-containing protein (PatX). Mol Microbiol 110:931-954.
If HetR and PatS (supplemented with HetN) are central to the mechanism by which heterocyst spacing is achieved (as previously suggested), then one might expect to find these proteins in all cyanobacteria that make spaced heterocysts. However, this is not the case (Figure 2). HetN is quite rare, and PatS is missing from a large family of heterocyst-producing cyanobacteria. HetR is indeed found in all heterocyst-producing cyanobacteria, but it is also present in multicellular cyanobacteria that lack heterocysts. These findings prompted us to look for a third RGSGR-containing protein, besides PatS and HetN.
What came from this search was a gene, patX, that is genetically linked to hetR and is found in those cyanobacteria where hetR is found, including those where PatS is absent and including the multicellular cyanobacteria that don't make heterocysts. The gene encodes a protein that has two conserved regions: an RGSGR motif and, like HetN, an N-terminal region that may determine a membrane anchor. It is regulated similarly to patS. Coupling the presence of HetR and PatX with the cyanobacterial phylogenetic tree, it is evident that the HetR/PatX regulatory pair long precedes the appearance of PatS and heterocyst differentiation and is strictly associated with multicellularity. It is not yet clear what role the pair plays in multicellular cyanobacteria.
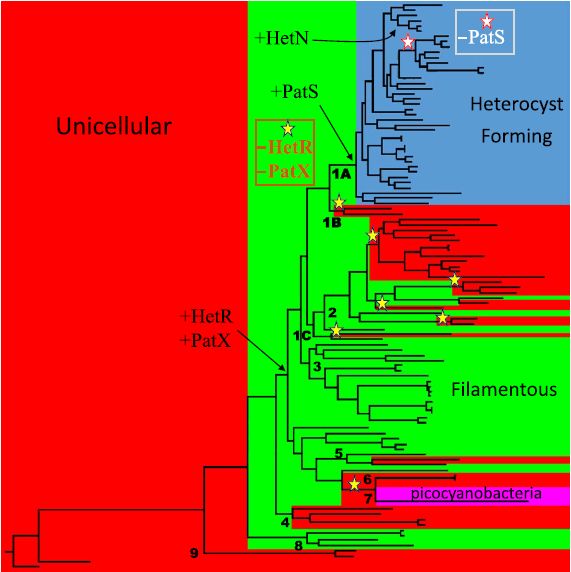
Figure 2. See text for explanation. Taken from Elhai & Khudyakov (2018).